








Formation, function, and replacement of membrane lipids in rhizobia
Principal investigator: Otto Geiger
Most bacteria possess phosphatidylglycerol (PG), cardiolipin (CL), phosphatidylethanolamine (PE) as major membrane lipids. However, some bacteria can also form the methylated derivatives of PE, monomethyl-PE (MMPE), dimethyl-PE (DMPE), and phosphatidylcholine (PC). In the bacterium Sinorhizobium meliloti , which can form nitrogen-fixing root nodules in symbiosis with the leguminous plant Medicago , PC can be formed by two distinct pathways. In the methylation pathway, PC is formed by three successive methylations of PE via the intermediates MMPE and DMPE using the methyl donor S -adenosyl-L-methionine, reactions catalyzed by one or several phospholipid N -methyltransferase(s) (Pmt). In the phosphatidylcholine synthase (Pcs) pathway, Pcs catalyzes the condensation of choline with CDP-diacylglycerol yielding PC and CMP as the products of this reaction ( Fig. 1 ). A mutagenic analysis of the pcs gene has allowed us to define which amino acid residues are essential for the function of this enzyme. Also, we have purified Pcs to near homogeneity and after crystallization of the enzyme we hope to obtain detailed structural information of this protein. This knowledge should permit us to define the active site of Pcs and to develop structure-based inhibitors for Pcs, which in cases where this enzyme is essential for the survival or pathogenesis of a bacterium ( Borrelia burgdorferi or Legionella pneumophila ) might function as antibiotics.
Mutants of S. meliloti deficient in pmt or pcs still produce wild type levels of PC and show no detectable phenotype. Double mutants, however, deficient in pmt and pcs are unable to form PC, cannot form nitrogen-fixing root nodules with their legume hosts, and show reduced growth when compared to the wild type. In medium containing tryptone/yeast extract (TY), PC-deficient sinorhizobial mutants do not grow at all. However, we could isolate PC-deficient suppressor mutants that regained growth on TY medium and presently we are mapping the mutation responsible for this growth suppression phenotype. By mapping such mutations, we would expect to identify genes the products of which are in some way controlled by PC. A control of these gene products by PC would define a specific molecular function for this lipid in S. meliloti .
Under phosphate-limiting conditions of growth, some eubacteria replace their membrane phospholipids by lipids not containing any phosphorus. In S. meliloti, these phosphorus-free lipids are sulfoquinovosyl diacylglycerol (SL), ornithine-containing lipids (OL), and diacylglyceryl N,N,N - trimethylhomoserine (DGTS) ( Fig. 2 ). Pulse/chase experiments indicate that during this replacement, phospholipids are subject to turnover and that they function as biosynthetic precursors for some of the phosphorus-free membrane lipids, namely of DGTS. It is thought that under phosphorus limitation, phospholipids are degraded to diacylglycerol which in turn serves as substrate in the first reaction of DGTS biosynthesis. We are presently identifying the gene and enzymatic activity responsible for phospholipid degradation under phosphorus-limiting conditions.
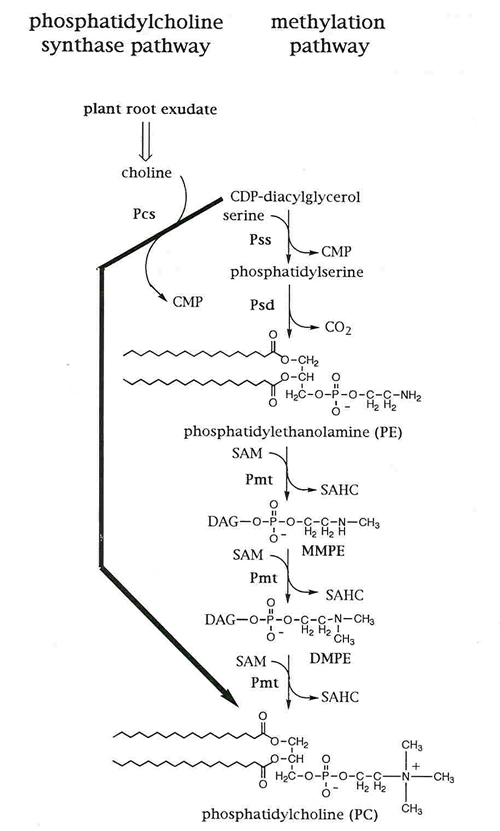
|
|
Fig. 1. Phosphatidylcholine (PC) biosynthesis in Sinorhizobium meliloti (taken from de Rudder et al . 1999. J. Biol. Chem. 274:20011-20016). SAM: S -adenosylmethionine; SAHC: S -adenosylhomocysteine; Psd: phosphatidylserine decarboxylase.
|